Single-cell technologies have transformed our ability to observe biology at unprecedented resolution. We can distinguish rare subpopulations, map heterogeneity, and characterise tissue complexity in exquisite detail, often in ways that were impossible only a few years ago. These advances have fundamentally reshaped discovery workflows, enabling new insights in target identification, patient stratification, and mechanism-of-action studies.
Yet a critical gap remains: much of what we infer about what cells do (their actual functional behaviour) still relies on proxies rather than direct measurement. In many cases, we are increasingly precise about what cells resemble, without equivalent certainty about what they are doing.
To explore why function has become a bottleneck in modern discovery, download our whitepaper: Advancing Drug Discovery with Function-Focused Profiling at the Single-Cell Level.
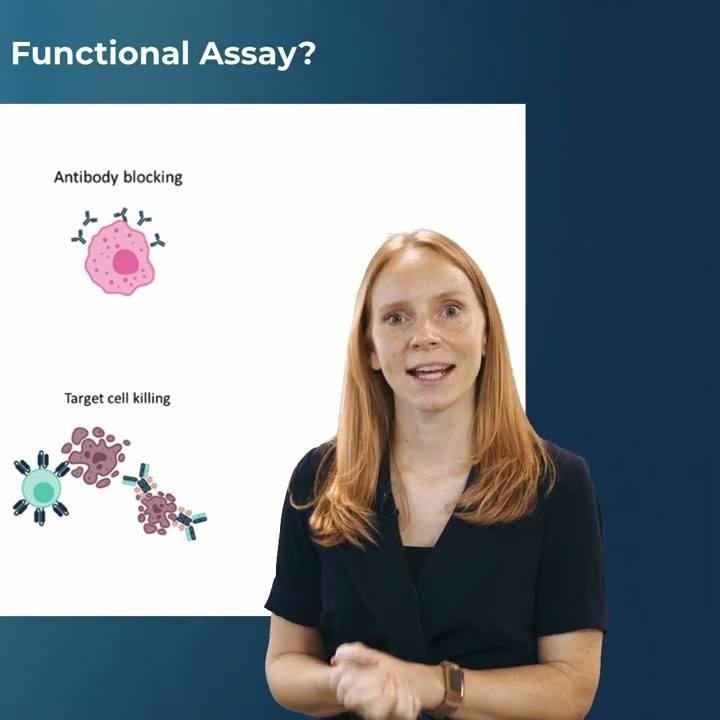
This matters because therapeutic success is determined not by molecular signatures alone, but by their functional consequences: activation, cytotoxicity, persistence, suppression, differentiation — or failure. These behaviours define whether a therapy works, how durable the response will be, and whether the mechanism translates beyond controlled experiments into patient biology.
Increasingly, our methods tell us which cells express the “right” markers or cluster into the “right” states — but not whether they execute the biology we need. And in drug discovery, that distinction is key. It shapes early selection decisions, determines which candidates advance, and influences whether efficacy risk is identified early or deferred into later, more expensive stages.
So if we can profile biology at single-cell resolution, why are we still guessing at function?
Resolution without understanding
A defining strength of single-cell analysis is its ability to resolve complexity. Technologies such as scRNA-seq allow us to dissect biological systems with remarkable precision, revealing structures and patterns that bulk methods inevitably obscure.
But resolution is not the same as understanding.
In many workflows, function is inferred from indirect signals: gene expression programs, surface marker panels, clustering patterns, pathway enrichment, and computational predictions. These approaches are powerful and have advanced the field significantly. But they rarely measure behaviour itself. Instead, they provide signatures that we interpret as functional states, often based on correlations established in specific contexts, cohorts, or experimental systems.
For years, this trade-off was unavoidable. Biology is difficult to measure at scale, and single-cell genomics offered a practical route to classification and comparison. But as datasets have grown and analytical sophistication has increased, the limitation has not disappeared. Transcriptional identity is not equivalent to functional behaviour, especially under perturbation, and when therapies, disease pressure, or microenvironmental constraints reshape cellular outputs in ways that are not captured cleanly by state labels.
The result is a subtle but persistent translation risk: we can generate large volumes of molecular data while still lacking confidence about the mechanisms that matter most for therapeutic performance.
Download the whitepaper for a deeper look at how inference shapes decision-making—and where direct functional measurement can shift risk earlier.
The inference problem at the heart of single-cell analysis
Single-cell research emerged to answer questions bulk measurements could not: which cells drive disease, how therapies reshape cellular landscapes, and why responses diverge between patients. These are functional questions — questions about behaviour under defined conditions.
Yet many of our most widely used readouts remain descriptive. We label cells “activated,” “exhausted,” “tumour-reactive,” or “effector-like” based on correlates, but correlation is not function. Cells with similar transcriptomic profiles can behave very differently, and similar behaviours can arise through distinct molecular routes, particularly under stress, inflammatory signalling, nutrient limitation, or therapeutic pressure.
This is the inference problem of single-cell analysis: we treat molecular patterns as functional truth, often without directly measuring the behaviours we care about.
That has practical consequences. When functional confirmation arrives only later — through secondary assays, downstream validation, or in vivo studies — the pipeline absorbs avoidable cost and delay. Candidates advance based on molecular promise but fail when tested on performance. Conversely, rare high-performing candidates may be missed if their function does not align cleanly with expected signatures.
At a time when discovery increasingly focuses on modalities where function is inseparable from mechanism, such as biologics, antibody engineering, and cell therapies, this gap becomes even more limiting. In these settings, the question is not only what state is present, but whether the therapeutic system reliably produces the necessary biological effect.
Bring function into focus
Single-cell profiling has given us extraordinary visibility into cellular diversity, but visibility alone does not guarantee translation. When functional behaviour is inferred rather than measured, efficacy risk is often discovered late — and after candidates have already accumulated cost, complexity, and momentum. As drug modalities become more function-dependent, the ability to directly quantify behaviour at single-cell resolution becomes less of a “nice to have” and more of a requirement for confident decision-making. The next step for single-cell discovery is not just higher resolution — it is functional certainty.